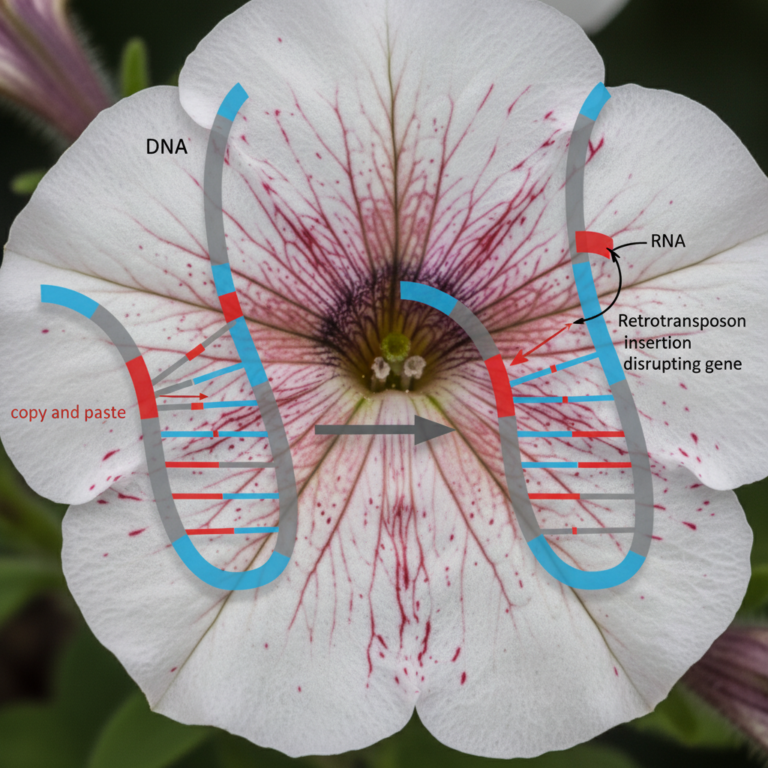
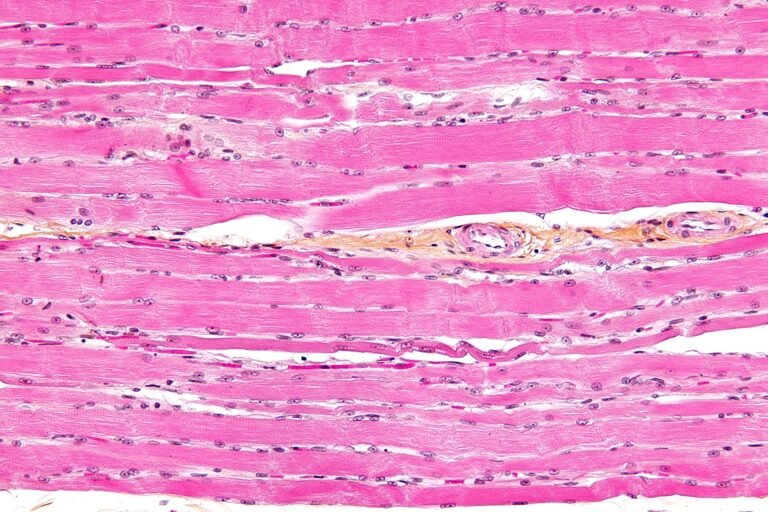
Advanced Glycation End Products
I have occasionally complained about my mother in law’s bland cooking methods. Boiled this, steamed that. How about a sauté? How about frying something? And yet, she is aging gracefully and the scientific evidence has begun to justify her insipid cooking. A growing body of research shows that advanced glycation end products (AGEs) sit at…